Species predicted to react based on 100% sequence homology:D. melanogaster, Xenopus, Zebrafish, Horse
Specificity / Sensitivity
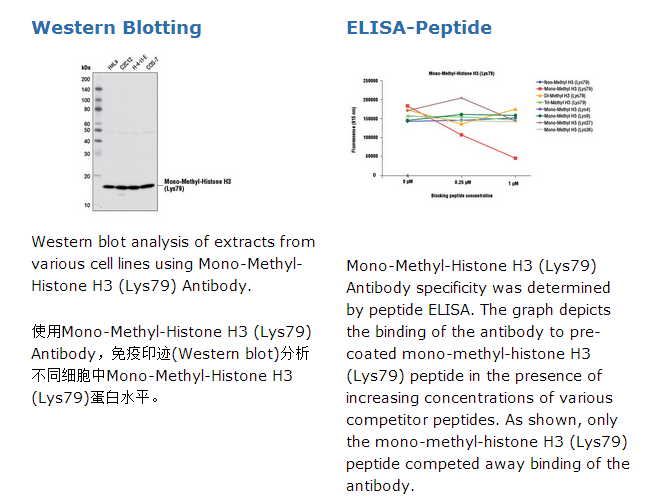
Mono-Methyl-Histone H3 (Lys79) Antibody recognizes endogenous levels of histone H3 protein only when mono-methylated at Lys79. The antibody does not cross-react with non-methylated, di-methylated, or tri-methylated histone H3 Lys79. In addition, the antibody does not cross-react with histone H3 mono-methylated at Lys4, Lys9, Lys27, or Lys36.
Source / Purification
Polyclonal antibodies are produced by immunizing animals with a synthetic peptide corresponding to residues surrounding mono-methyl Lys79 of human histone H3 protein. Antibodies are purified by protein A and peptide affinity chromatography.
Background
The nucleosome, made up of four core histone proteins (H2A, H2B, H3, and H4), is the primary building block of chromatin. Originally thought to function as a static scaffold for DNA packaging, histones have now been shown to be dynamic proteins, undergoing multiple types of post-translational modifications, including acetylation, phosphorylation, methylation, and ubiquitination (1). Histone methylation is a major determinant for the formation of active and inactive regions of the genome and is crucial for the proper programming of the genome during development (2,3). Arginine methylation of histones H3 (Arg2, 17, 26) and H4 (Arg3) promotes transcriptional activation and is mediated by a family of protein arginine methyltransferases (PRMTs), including the co-activators PRMT1 and CARM1 (PRMT4) (4). In contrast, a more diverse set of histone lysine methyltransferases has been identified, all but one of which contain a conserved catalytic SET domain originally identified in the Drosophila Su(var)3-9, Enhancer of zeste, and Trithorax proteins. Lysine methylation occurs primarily on histones H3 (Lys4, 9, 27, 36, 79) and H4 (Lys20) and has been implicated in both transcriptional activation and silencing (4). Methylation of these lysine residues coordinates the recruitment of chromatin modifying enzymes containing methyl-lysine binding modules such as chromodomains (HP1, PRC1), PHD fingers (BPTF, ING2), tudor domains (53BP1), and WD-40 domains (WDR5) (5-8). The discovery of histone demethylases such as PADI4, LSD1, JMJD1, JMJD2, and JHDM1 has shown that methylation is a reversible epigenetic marker (9).



 京公网安备11010802025653 版权所有:北京逸优科技有限公司
京公网安备11010802025653 版权所有:北京逸优科技有限公司
