Phospho-Aurora A (Thr288)/Aurora B (Thr232)/Aurora C (Thr198) (D13A11) XP® Rabbit mAb (Alexa Fluor® 488 Conjugate)能够分别识别内源性苏氨酸(288位)、苏氨酸(232位)、苏氨酸(198位)磷酸化的Aurora A/B/C蛋白。该单克隆抗体是由合成的人源的针对Aurora B蛋白苏氨酸(232位)的肽段免疫动物生产的。该抗体经过Alexa Fluor® 488荧光染料标记并且使用人类细胞采用直接流式细胞术和免疫荧光分析通过了内部测试。该抗体预计于未标记的Phospho-Aurora A (Thr288)/Aurora B (Thr232)/Aurora C (Thr198) (D13A11) XP® Rabbit mAb #2914 抗体具有同样的种属交叉反应性。极光激酶属于有丝分裂过程中高度保守的丝氨酸/苏氨酸激酶家族,在哺乳动物中已确定了三个成员:极光激酶A,极光激酶B和极光激酶C(1,2)。时空表达模式和有丝分裂细胞中极光激酶亚细胞定位的研究表明其和有丝分裂结构的关系。极光激酶功能的影响跨越G2期和细胞分裂期并可能涉及到关键的细胞周期事件,如中心体复制、染色体双向定位和隔离、卵裂沟定位和内移(3)。极光激酶A可以在中心体(沿纺锤体微管方向)和有丝分裂增殖细胞的胞质中被检测到。极光激酶A蛋白水平在G1和S期较低而在细胞周期的G2/M期达峰值。极光激酶A催化结构域苏氨酸(288位)的磷酸化能够增强激酶的活性。极光激酶A参与中心体分离、成熟和纺锤体组装及稳定。极光激酶B的蛋白表达同样在细胞周期的G2/M期达峰值; 极光激酶B的激酶活性在分裂中期到有丝分裂结束的过渡阶段最高。在重新定位到有丝分裂后期纺锤体之前,极光激酶B和分裂前期的染色体密切相关。极光激酶B通过控制微管-着丝粒附着和细胞质分裂调节染色体分离。G2/M期过渡时极光激酶A和极光激酶B的表达紧密配合组蛋白H3的磷酸化(4,5); 多种人类癌症中都发现了这些激酶的过度表达(2,4)。极光激酶C集中于有丝分裂后期到细胞质分裂期的中心体中,而且其mRNA和蛋白质水平在G2/M期达峰值。虽然典型的极光激酶C的表达仅限于睾丸,但是在各种肿瘤细胞株中都可以检测到其过度表达(6)。
Phospho-Aurora A (Thr288)/Aurora B (Thr232)/Aurora C (Thr198) (D13A11) XP® Rabbit mAb (Alexa Fluor® 488 Conjugate) detects endogenous levels of Aurora A/B/C only when phosphorylated at either Thr288, Thr232 or Thr198 respectively.
Source / Purification
Monoclonal antibody is produced by immunizing animals with a synthetic peptide corresponding to residues surrounding Thr232 of human Aurora B.
Description
This Cell Signaling Technology antibody is conjugated to Alexa Fluor® 488 fluorescent dye and tested in-house for direct flow cytometric and immunofluorescent analysis in human cells. This antibody is expected to exhibit the same species cross-reactivity as the unconjugated Phospho-Aurora A (Thr288)/Aurora B (Thr232)/Aurora C (Thr198) (D13A11) XP® Rabbit mAb #2914.
Background
Aurora kinases belong to a highly conserved family of mitotic serine/threonine kinases with three members identified among mammals: Aurora A, B, and C (1,2). Studies on the temporal expression pattern and subcellular localization of Aurora kinases in mitotic cells suggest an association with mitotic structure. Their functional influences span from G2 phase to cytokinesis and may be involved in key cell cycle events such as centrosome duplication, chromosome bi-orientation and segregation, cleavage furrow positioning, and ingression (3). Aurora A is detected at the centrosomes, along mitotic spindle microtubules, and in the cytoplasm of mitotically proliferating cells. Aurora A protein levels are low during G1 and S phases and peak during the G2/M phase of the cell cycle. Phosphorylation of Aurora A at Thr288 in its catalytic domain increases kinase activity. Aurora A is involved in centrosome separation, maturation, and spindle assembly and stability. Expression of Aurora B protein also peaks during the G2/M phase of the cell cycle; Aurora B kinase activity peaks at the transition from metaphase to the end of mitosis. Aurora B associates with chromosomes during prophase prior to relocalizing to the spindle at anaphase. Aurora B regulates chromosome segregation through the control of microtubule-kinetochore attachment and cytokinesis. Expression of both Aurora A and Aurora B during the G2/M phase transition is tightly coordinated with histone H3 phosphorylation (4,5); overexpression of these kinases is seen in a variety of human cancers (2,4). Aurora C localizes to the centrosome from anaphase to cytokinesis and both mRNA and protein levels peak during G2/M phase. Although typical Aurora C expression is limited to the testis, overexpression of Aurora C is detected in various cancer cell lines (6).



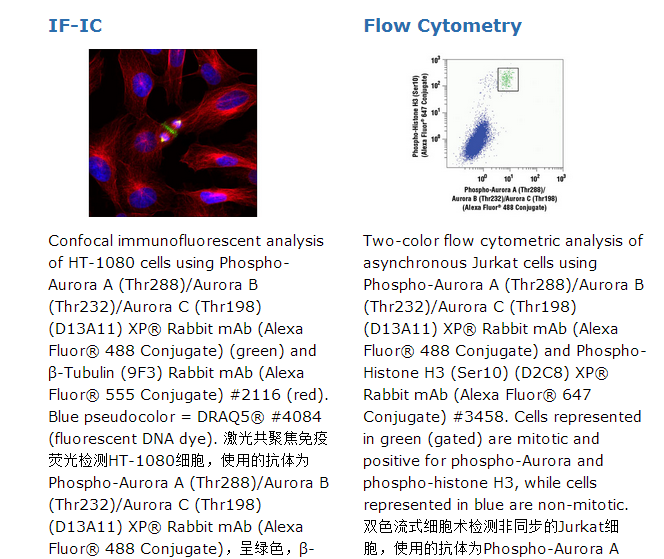
 京公网安备11010802025653 版权所有:北京逸优科技有限公司
京公网安备11010802025653 版权所有:北京逸优科技有限公司
