Cell Signaling Technology (CST)公司的SignalSilence® CHD1 siRNA II可以帮助研究者通过RNA干扰特异性地抑制CHD1蛋白表达,这种方法可以通过将双链RNA分子传递到细胞内从而使基因表达有选择的沉默。来自CST的所有的SignalSilence®siRNA产品都是经过内部严格检测的,并且通过Western blot 分析证明确实能够减少目的蛋白的表达。通过三苯甲基分析每个碱基以监测寡核苷酸的合成,确保合适的配对效率。随后寡核苷酸通过亲和固相萃取法纯化。退火的RNA双链通过质谱分析来证实其精确的组成。每一批产品都通过质谱分析与前面的产品进行比较,来保证不同批次之间的最大一致性。Chromodomain-helicase-DNA-binding domain (CHD)蛋白已经在多种生物中被鉴定(1,2)。由九个成员组成的蛋白家族已经被分为三个亚家族:subfamily I (CHD1和CHD2)、subfamily II (CHD3和CHD4)和subfamily III (CHD5、CHD6、CHD7、CHD8、CHD9)。所有CHD蛋白包含两个串联的N端chromodomains、一个SWI/SNF相关的ATPase结构域和一个C端DNA结合区域 (1,2)。chromodomains有助于结合到组蛋白的已甲基化赖氨酸残基,并且可与染色质的特异区域相互作用。SWI/SNF相关的ATPase区域可利用来自ATP水解的能量去修饰染色质结构。CHD1是一个常染色质蛋白,它与活化基因的启动子有关联,并且该蛋白对于在胚胎干细胞中染色质开放性和多能性的维持是需要的(3-6)。CHD1蛋白的两个chromodomains有助于它到活化基因的招募,该过程通过结合到histone H3的methyl-lysine 4位点,它是一个与转录激活相关联的标记物(4-6)。酵母CHD1蛋白是SAGA和SLIK组蛋白乙酰化转移酶复合物的一个成分,并且被认为在转录激活中联系着组蛋白甲基化和组蛋白乙酰化作用(6)。CHD2蛋白不具有明显特征;然而,小鼠基因敲除研究认为在发育和肿瘤抑制中有重要作用。纯合子CHD2敲除老鼠显示延迟生长和围产期致死(7)。杂合子敲除老鼠显示除了增加的髓外造血和易感性淋巴瘤之外,还出现死亡率增加和所有器官不正常(7,8)。CHD2突变细胞在造血干细胞分化中缺失,并且展示异常的DNA损伤反应(8)。
SignalSilence® CHD1 siRNA II from Cell Signaling Technology (CST) allows the researcher to specifically inhibit CHD1 expression using RNA interference, a method whereby gene expression can be selectively silenced through the delivery of double stranded RNA molecules into the cell. All SignalSilence® siRNA products from CST are rigorously tested in-house and have been shown to reduce target protein expression by western analysis.
Quality Control
Oligonucleotide synthesis is monitored base by base through trityl analysis to ensure appropriate coupling efficiency. The oligo is subsequently purified by affinity-solid phase extraction. The annealed RNA duplex is further analyzed by mass spectrometry to verify the exact composition of the duplex. Each lot is compared to the previous lot by mass spectrometry to ensure maximum lot-to-lot consistency.
Directions for Use
CST recommends transfection with 100 nM SignalSilence® CHD1 siRNA II 48 to 72 hours prior to cell lysis. For transfection procedure, follow protocol provided by the transfection reagent manufacturer. Please feel free to contact CST with any questions on use.
Background
Chromodomain-helicase-DNA-binding domain (CHD) proteins have been identified in a variety of organisms (1,2). This family of proteins, which consists of nine members, has been divided into three separate subfamilies: subfamily I (CHD1 and CHD2), subfamily II (CHD3 and CHD4), and subfamily III (CHD5, CHD6, CHD7, CHD8, and CHD9). All of the CHD proteins contain two tandem N-terminal chromodomains, a SWI/SNF-related ATPase domain, and a C-terminal DNA binding domain (1,2). The chromodomains facilitate binding to methylated lysine residues of histone proteins and confer interactions with specific regions of chromatin. The SWI/SNF-related ATPase domain utilizes the energy from ATP hydrolysis to modify chromatin structure. CHD1 is a euchromatic protein that associates with the promoters of active genes, and is required for the maintenance of open chromatin and pluripotency in embryonic stem cells (3-6). The two chromodomains of CHD1 facilitate its recruitment to active genes by binding to methyl-lysine 4 of histone H3, a mark associated with transcriptional activation (4-6). Yeast CHD1 is a component of the SAGA and SLIK histone acetyltransferase complexes, and is believed to link histone methylation with histone acetylation during transcriptional activation (6). The CHD2 protein is not well characterized; however, mouse knockout studies suggest important functions in development and tumor suppression. Homozygous CHD2 knockout mice exhibit delayed growth and perinatal lethality (7). Heterozygous knockout mice show increased mortality and gross organ abnormalities, in addition to increased extramedullary hematopoiesis and susceptibility to lymphomas (7,8). CHD2 mutant cells are defective in hematopoietic stem cell differentiation and exhibit aberrant DNA damage responses (8).



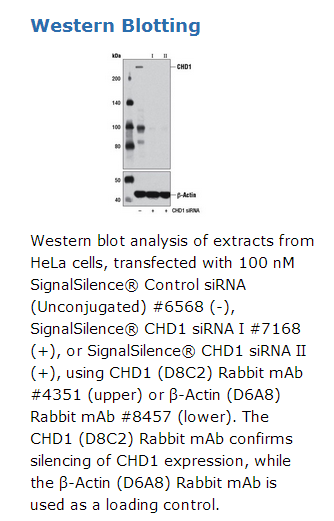
 京公网安备11010802025653 版权所有:北京逸优科技有限公司
京公网安备11010802025653 版权所有:北京逸优科技有限公司
