Species predicted to react based on 100% sequence homology:Monkey
Description
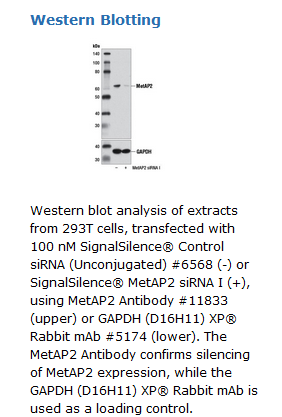
SignalSilence® MetAP2 siRNA I from Cell Signaling Technology (CST) allows the researcher to specifically inhibit MetAP2 expression using RNA interference, a method whereby gene expression can be selectively silenced through the delivery of double stranded RNA molecules into the cell. All SignalSilence® siRNA products from CST are rigorously tested in-house and have been shown to reduce target protein expression by western analysis.
Quality Control
Oligonucleotide synthesis is monitored base by base through trityl analysis to ensure appropriate coupling efficiency. The oligo is subsequently purified by affinity-solid phase extraction. The annealed RNA duplex is further analyzed by mass spectrometry to verify the exact composition of the duplex. Each lot is compared to the previous lot by mass spectrometry to ensure maximum lot-to-lot consistency.
Directions for Use
CST recommends transfection with 100 nM SignalSilence® MetAP2 siRNA I 48 to 72 hours prior to cell lysis. For transfection procedure, follow protocol provided by the transfection reagent manufacturer. Please feel free to contact CST with any questions on use. Each vial contains the equivalent of 100 transfections, which corresponds to a final siRNA concentration of 100 nM per transfection in a 24-well plate with a total volume of 300 μl per well.
Background
Eukaryotic initiation factor 2 (eIF2)-associated glycoprotein, p67/methionine aminopeptidase 2 (MetAP2) is one of the three known MetAPs responsible for the co-translational processing of the N-terminal initiator methionine from nascent proteins in cells. MetAP2 regulates the rates of global protein synthesis by controlling the levels of eIF2α phosphorylation (1). MetAP2 has also been shown to bind Erk1/2 to inhibit their activation and activity, thus connecting the protein synthesis machinery with the cell signaling pathway mediated by Erk1/2 MAP kinases (2-4). Although MetAP2 is characterized as having aminopeptidase activity that removes the N-terminal methionine from nascent peptides in vitro, mounting evidence suggests that MetAP2 has no methionine aminopeptidase activity. Rather, MetAP2 possesses auto-proteolytic activity that can be inhibited by several small molecule inhibitors including anti-angiogenic drugs, fumagillin and its derivatives (5). It has also been demonstrated that O-GlcNAcylation of MetAP2 plays a major role in its stability, eIF2α binding, and maintenance of eIF2α phosphorylation (6). MetAP2 knockout mice show embryonic lethality, suggesting its role in embryonic development and survival at the initiation of gastrulation (7). It is likely that lowering the levels of MetAP2 in mammalian cells causes cell growth inhibition and leads to apoptosis due to the high levels of eIF2α phosphorylation that inhibits global protein synthesis (8). During pathological or various stress conditions, MetAP2 dissociates from eIF2 subunits possibly due to its deglycosylation-induced autoproteolytic cleavage. As a result, eIF2α becomes hyperphosphorylated and global protein synthesis is inhibited. eIF2 complex-dissociated MetAP2 also displays a higher affinity toward Erk1/2, which results in the blockade of Erk1/2 activity. Thus, MetAP2 mediates cooperation between cell signaling and protein synthesis machinery to regulate cell growth and proliferation during physiological and pathological conditions (9). Research studies have shown higher expression of MetAP2 in human cancers, supporting the contention that MetAP2 plays a role in oncogenesis. For example, investigators have reported high MetAP2 expression in follicular lymphomas, large B-cell lymphomas, and Burkitt's lymphomas (10). Elevated expression of MetAP2 has also been reported in human colorectal adenocarcinomas (11).



 京公网安备11010802025653 版权所有:北京逸优科技有限公司
京公网安备11010802025653 版权所有:北京逸优科技有限公司
