SignalSilence® PLK1 siRNA II 可以抑制人和猴的PLK1蛋白表达。Cell Signaling Technology (CST) 公司的SignalSilence® PLK1 siRNA 帮助研究人员使用RNA干扰技术特异性抑制PLK1表达,该技术通过向细胞内导入双链RNA分子以选择性的抑制某些基因的表达。所有CST公司的SignalSilence® siRNA都经过了严格的内部测试,western分析显示能够能够降低目标蛋白的表达。寡核苷酸合成经过了三苯甲基的逐碱基分析以确保耦合效率。随后经过亲和固相萃取纯化。退火的RNA双链在经过质谱分析以确保双链的准确性。新批次都会经过质谱分析与之前的批次进行比对以保证不同批次之间的稳定性。CST建议转染100 nM SignalSilence® PLK1 siRNA II,48-72小时后裂解细胞。转染的步骤请参阅转染试剂的说明书。关于使用问题,可以随时咨询CST。 每管试剂能进行100次转染,该转染次数以总体积为300μl 的1个24孔板孔中siRNA终浓度100 nM来进行计算。哺乳动物细胞中有至少4中不同的polo-样激酶:PLK1,PLK2,PLK3和PLK4/SAK(1). PLK1在有丝分裂过程中发挥了众多功能,尤其是在调控有丝分裂的开始和退出。Cdc25去磷酸化cdc2 (Thr14/Tyr15)能够激活有丝分裂促进因子(MPF),cdc2/cyclin B1。PLK1磷酸化cdc25C的198位丝氨酸和细胞周期蛋白B1的133位丝氨酸会促使这些蛋白从细胞质进入细胞核(2-5)。PLK1磷酸化Myt1的426位丝氨酸和495苏氨酸将失活Myt1,Myt1是一个激酶能够磷酸化cdc2的14位苏氨酸/15位酪氨酸(6)。Polo-样激酶还可以磷酸化黏连蛋白亚基SCC1,使黏连蛋白从染色体臂端移到着丝粒(7)。有丝分裂结束时需要激活细胞分裂后期促进复合物(APC)(8),APC是一个泛素连接酶,作用包括从着丝粒上移除黏连蛋白, 降解degradation of securin, cyclin A, cyclin B1, Aurora A, 和cdc20(9)。体外实验已经证明PLK1可以磷酸化APC亚基Apc1, cdc16, 和cdc27,这一机制据信调控了有丝分裂期结束(10,11)。用天冬氨酸替换PLK1的210位苏氨酸能够提高其及酶活性,并延迟/阻断细胞进入有丝分裂,而Ser137Asp替换则能引发细胞处于s期阻滞(12)。此外,研究发现DNA损伤会抑制PLK1的及酶活性,Thr210Asp突变则能抵御这一抑制效果(13)。体内试验报道有丝分裂时PLK1的137位丝氨酸和210位苏氨酸发生磷酸化;DNA损伤则会阻止这些位点的磷酸化(14)。
原厂资料:
Homology
Species predicted to react based on 100% sequence homology:Monkey
Specificity / Sensitivity
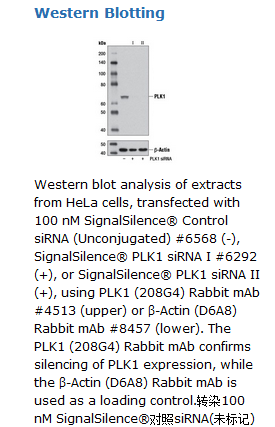
SignalSilence® PLK1 siRNA II inhibits human and monkey PLK1 expression.
Description
SignalSilence® PLK1 siRNA II from Cell Signaling Technology (CST) allows the researcher to specifically inhibit PLK1 expression using RNA interference, a method whereby gene expression can be selectively silenced through the delivery of double stranded RNA molecules into the cell. All SignalSilence® siRNA products from CST are rigorously tested in-house and have been shown to reduce target protein expression by western analysis.
Quality Control
Oligonucleotide synthesis is monitored base by base through trityl analysis to ensure appropriate coupling efficiency. The oligo is subsequently purified by affinity-solid phase extraction. The annealed RNA duplex is further analyzed by mass spectrometry to verify the exact composition of the duplex. Each lot is compared to the previous lot by mass spectrometry to ensure maximum lot-to-lot consistency.
Quality Control
Oligonucleotide synthesis is monitored base by base through trityl analysis to ensure appropriate coupling efficiency. The oligo is subsequently purified by affinity-solid phase extraction. The annealed RNA duplex is further analyzed by mass spectrometry to verify the exact composition of the duplex. Each lot is compared to the previous lot by mass spectrometry to ensure maximum lot-to-lot consistency.
Background
At least four distinct polo-like kinases exist in mammalian cells: PLK1, PLK2, PLK3, and PLK4/SAK (1). PLK1 apparently plays many roles during mitosis, particularly in regulating mitotic entry and exit. The mitosis promoting factor (MPF), cdc2/cyclin B1, is activated by dephosphorylation of cdc2 (Thr14/Tyr15) by cdc25C. PLK1 phosphorylates cdc25C at Ser198 and cyclin B1 at Ser133 causing translocation of these proteins from the cytoplasm to the nucleus (2-5). PLK1 phosphorylation of Myt1 at Ser426 and Thr495 has been proposed to inactivate Myt1, one of the kinases known to phosphorylate cdc2 at Thr14/Tyr15 (6). Polo-like kinases also phosphorylate the cohesin subunit SCC1, causing cohesin displacement from chromosome arms that allow for proper cohesin localization to centromeres (7). Mitotic exit requires activation of the anaphase promoting complex (APC) (8), a ubiquitin ligase responsible for removal of cohesin at centromeres, and degradation of securin, cyclin A, cyclin B1, Aurora A, and cdc20 (9). PLK1 phosphorylation of the APC subunits Apc1, cdc16, and cdc27 has been demonstrated in vitro and has been proposed as a mechanism by which mitotic exit is regulated (10,11). Substitution of Thr210 with Asp has been reported to elevate PLK1 kinase activity and delay/arrest cells in mitosis, while a Ser137Asp substitution leads to S-phase arrest (12). In addition, while DNA damage has been found to inhibit PLK1 kinase activity, the Thr210Asp mutant is resistant to this inhibition (13). PLK1 has been reported to be phosphorylated in vivo at Ser137 and Thr210 in mitosis; DNA damage prevents phosphorylation at these sites (14).
注意事项:
Storage: PLK1 siRNA II is supplied in RNAse-free water. Aliquot
and store at -20ºC



 京公网安备11010802025653 版权所有:北京逸优科技有限公司
京公网安备11010802025653 版权所有:北京逸优科技有限公司
